
Super Flu Outbreak: Not Merely a Virus, but an Alarm of the Body’s Ecosystem
The recent global surge of severe influenza cases, popularly termed “Super Flu,” has been dominated by Influenza A H3N2, including emerging subclades such as Subclade K, which demonstrate enhanced transmissibility, immune escape, and clinical severity.
JAKARTA – The recent global surge of severe influenza cases, popularly termed “Super Flu,” has been dominated by Influenza A H3N2, including emerging subclades such as Subclade K, which demonstrate enhanced transmissibility, immune escape, and clinical severity.
While virological evolution contributes to this phenomenon, accumulating evidence indicates that host biological vulnerability plays a decisive role in determining disease severity. This article proposes that the current Super Flu wave should be interpreted not merely as a viral outbreak, but as an alarm of disrupted host ecosystem integrity.
Influenza A H3N2 is an enveloped virus whose infectivity critically depends on the integrity of its lipid membrane. In parallel, modern populations exhibit widespread gut dysbiosis, chronic low-grade inflammation, impaired mucosal barriers, and reduced immunological adaptability, largely driven by ultra-processed diets, low fiber intake, metabolic stress, and environmental exposures.
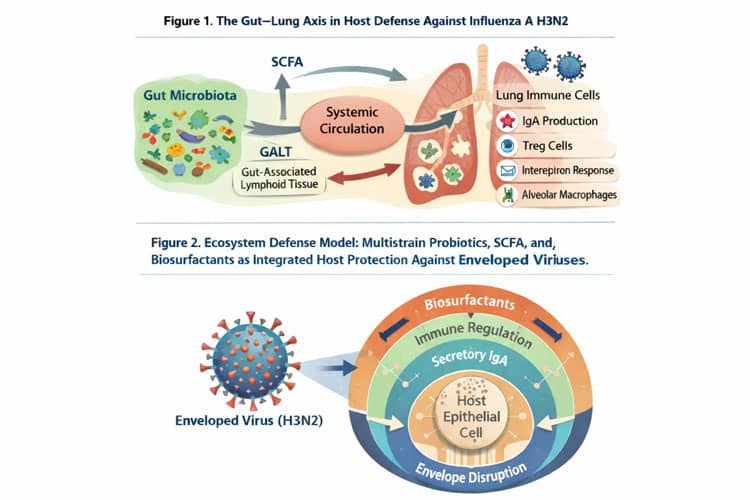
These conditions compromise the Gut–Lung Axis, weaken mucosal immunity, reduce secretory IgA production, impair interferon responses, and increase epithelial permeability, thereby creating a permissive biological environment for viral invasion and propagation.
This review integrates current knowledge on the role of multistrain probiotics, short-chain fatty acids (SCFA), and microbially derived biosurfactants as ecosystem-level biological defenses against enveloped respiratory viruses.
Multistrain probiotics restore microbial diversity and functional redundancy, enhance SCFA production, strengthen epithelial tight junctions, and recalibrate immune responses. SCFA act as key metabolic and epigenetic regulators of mucosal integrity, antiviral immunity, and inflammatory resolution.
Furthermore, probiotic-derived biosurfactants possess amphipathic properties capable of destabilizing lipid viral envelopes, providing a mechanistically plausible “biological envelope disruption” strategy against H3N2.
Collectively, this article argues for a paradigm shift from pathogen-centered interventions toward host ecosystem restoration as a foundational strategy for influenza prevention and severity mitigation. Super Flu is framed as a manifestation of ecosystem failure rather than viral exceptionalism.
Rebuilding gut eubiosis, mucosal integrity, and metabolic resilience through multistrain probiotics, SCFA support, and dietary modulation represents a rational, sustainable, and causally coherent approach to strengthening host defenses against current and future influenza threats.
Introduction: Super Flu as a Global and National Alarm
The recent global surge of severe influenza cases, popularly termed “Super Flu,” has been dominated by Influenza A H3N2, including emerging subclades such as Subclade K, which demonstrate enhanced transmissibility, immune escape, and clinical severity (Petrova & Russell, 2018; Skowronski et al., 2019).
Surveillance data from multiple regions indicate increased hospitalization rates and prolonged disease courses compared to previous influenza seasons (Belongia et al., 2016).
Indonesia mirrors this global trend, with a marked increase in influenza-like illness (ILI) cases presenting with high-grade fever, severe myalgia, prolonged fatigue, and delayed recovery. Importantly, a significant proportion of these cases occur in individuals without classical comorbidities, suggesting that host biological vulnerability, rather than viral virulence alone, plays a critical role in disease severity (Dang & Marsland, 2019).
These observations support a paradigm shift: Super Flu should not be interpreted solely as a virological phenomenon, but as a manifestation of disrupted host ecosystem integrity.
Characteristics of Influenza A H3N2 Subclade K and the Significance of Lipid Envelope
Influenza A H3N2 is recognized as the subtype with the highest rate of antigenic drift and immune escape, largely due to continuous mutations in hemagglutinin (HA) and neuraminidase (NA) proteins (Petrova & Russell, 2018). These mutations significantly reduce vaccine effectiveness across seasons (Belongia et al., 2016).
Structurally, H3N2 is an enveloped virus possessing a lipid bilayer envelope derived from the host cell membrane (Schoeman & Fielding, 2019). The integrity of this lipid envelope is essential for viral attachment, fusion, and entry into host epithelial cells (Otter et al., 2016). Enveloped viruses are inherently vulnerable to agents that disrupt lipid bilayers, including surfactants and biosurfactants (Kampf et al., 2020).
The biological similarity between H3N2 and SARS-CoV-2 as enveloped viruses highlights a shared structural vulnerability: disruption of the lipid envelope renders the virus non-infectious (Kampf et al., 2020).
Gut–Lung Axis: The Central Determinant of Respiratory Immunity
The Gut–Lung Axis represents a bidirectional communication network between intestinal microbiota and pulmonary immunity mediated by immune cells, cytokines, and microbial metabolites (Budden et al., 2017; Dang & Marsland, 2019). Experimental models demonstrate that germ-free or dysbiotic mice exhibit impaired immune responses to influenza infection (Ichinohe et al., 2011).
Microbial-derived metabolites, particularly short-chain fatty acids (SCFA), regulate hematopoiesis, macrophage function, and antiviral interferon responses in the lung (Trompette et al., 2014; Koh et al., 2016). This axis establishes that respiratory immunity is inseparable from gut ecosystem health.
Gut Dysbiosis, Mucosal Breakdown, and Immune Collapse
Gut dysbiosis is characterized by reduced microbial diversity, depletion of beneficial commensals, and overgrowth of pathobionts (Bischoff et al., 2014). This condition leads to increased intestinal permeability (“leaky gut”), allowing lipopolysaccharide (LPS) translocation into systemic circulation, triggering chronic low-grade inflammation (Cani et al., 2007).
Chronic endotoxemia induces immune exhaustion and impairs adaptive flexibility, resulting in a paradoxical state where the immune system is persistently activated but functionally inefficient (De Punder & Pruimboom, 2015). This explains why individuals without overt disease can still experience severe influenza manifestations.
Chronic Inflammation and Modern Diet as Accelerators of Vulnerability
Modern diets high in refined carbohydrates, ultra-processed foods, and low in dietary fiber directly promote dysbiosis and reduce SCFA production (Koh et al., 2016; Tan et al., 2014). Reduced SCFA availability compromises epithelial tight junction integrity, IgA secretion, and anti-inflammatory signaling (Furusawa et al., 2013).
Trompette et al. (2014) provided direct evidence that dietary fiber intake enhances SCFA production and confers protection against influenza by modulating immune cell function in the lung. This establishes diet as an immunological determinant, not merely a metabolic factor.
Hyperglycemia and protein glycation further impair immune receptor function and increase viral replication efficiency (Cani et al., 2007; De Punder & Pruimboom, 2015).
Multistrain Probiotics as Ecosystem Restorers
Multistrain probiotics restore microbial diversity and functional redundancy, mimicking natural ecosystem complexity (Hao et al., 2015). Meta-analyses demonstrate that probiotic supplementation reduces the incidence, duration, and severity of upper respiratory tract infections (Hao et al., 2015; King et al., 2014; Wang et al., 2016).
Mechanistically, probiotics enhance secretory IgA production, activate natural killer cells, and regulate Th1/Th2 balance (Kim et al., 2013). These effects are not strain-isolated but arise from inter-strain metabolic cooperation.
SCFA as Metabolic and Epigenetic Regulators of Immunity
SCFA, particularly butyrate, function as histone deacetylase inhibitors and promote differentiation of regulatory T cells (Furusawa et al., 2013). Atarashi et al. (2013) demonstrated that specific commensal clusters directly induce Treg expansion.
SCFA also enhance interferon responses and macrophage antiviral activity in the lung (Trompette et al., 2014). These mechanisms explain how gut-derived metabolites exert systemic antiviral effects.
Biosurfactants as Natural Antiviral Envelope Disruptors
Microbial biosurfactants are amphipathic molecules capable of disrupting lipid bilayers (Banat et al., 2010). Rodrigues et al. (2006) reported strong antimicrobial and anti-adhesive properties of biosurfactants, including activity against enveloped pathogens.
Kok & Nyotohadi (2024) demonstrated that multistrain probiotic extracts rich in biosurfactants significantly disrupted the lipid envelope of SARS-CoV-2 in vitro. Given the structural similarity between SARS-CoV-2 and H3N2 as enveloped viruses, this mechanism is directly relevant.
Unlike antiviral drugs targeting viral enzymes, biosurfactants act on physical membrane integrity, making them resistant to viral mutation escape (Banat et al., 2010).
Translational Evidence from COVID-19
Alterations in gut microbiota composition correlate with COVID-19 severity (Zuo et al., 2020). Systematic reviews indicate that probiotic supplementation may reduce inflammation and improve clinical outcomes in COVID-19 patients (Liu et al., 2021). These findings support the translational relevance of ecosystem-based interventions for other respiratory enveloped viruses, including H3N2.
Integrated Ecosystem Defense Model
Multistrain probiotics restore microbial balance, SCFA reinforce mucosal barriers and immune regulation, and biosurfactants disrupt viral envelopes. Together, these mechanisms form a multilayered ecosystem defense system (Budden et al., 2017; Trompette et al., 2014; Banat et al., 2010).
This model shifts the focus from pathogen elimination to host ecosystem restoration.
Conclusion
The current surge of severe influenza dominated by Influenza A H3N2, including Subclade K, cannot be adequately explained by viral evolution alone. This review demonstrates that host ecosystem vulnerability—characterized by gut dysbiosis, impaired mucosal barriers, chronic low-grade inflammation, and reduced immunological adaptability—is a decisive determinant of disease severity. As an enveloped virus, H3N2 critically depends on lipid membrane integrity and efficient mucosal entry, both of which are strongly influenced by the host’s biological terrain.
Multistrain probiotics, short-chain fatty acids (SCFA), and microbially derived biosurfactants collectively represent a coherent ecosystem-based defense strategy. Multistrain probiotics restore microbial diversity and functional redundancy, enhance SCFA production, and recalibrate immune responses. SCFA strengthen epithelial tight junctions, promote secretory IgA production, modulate interferon signaling, and facilitate inflammatory resolution. Biosurfactants, through their amphipathic properties, provide a mechanistically plausible biological envelope disruption mechanism against lipid-enveloped viruses such as H3N2.
These mechanisms act synergistically to fortify the Gut–Lung Axis, stabilize mucosal immunity, and eliminate permissive conditions for viral invasion and propagation. Accordingly, Super Flu should be reframed as an ecosystem failure rather than viral exceptionalism. Restoring host biological resilience through multistrain probiotic supplementation, SCFA support, and dietary fiber modulation constitutes a rational, sustainable, and causally coherent strategy to mitigate influenza severity and enhance population-level resistance to future respiratory viral threats.
References:
Belongia, E. A., Simpson, M. D., King, J. P., Sundaram, M. E., Kelley, N. S., Osterholm, M. T., & McLean, H. Q. (2016). Variable influenza vaccine effectiveness by subtype: A systematic review and meta-analysis. Clinical Infectious Diseases, 62(5), 584–594.
https://doi.org/10.1093/cid/civ1049
Petrova, V. N., & Russell, C. A. (2018). The evolution of seasonal influenza viruses. Nature Reviews Microbiology, 16(1), 47–60.
https://doi.org/10.1038/nrmicro.2017.118
Skowronski, D. M., Chambers, C., Sabaiduc, S., De Serres, G., Winter, A. L., Dickinson, J. A., … & Gubbay, J. B. (2019). Integrated sentinel surveillance linking genetic, antigenic, and epidemiologic monitoring of influenza virus. Euro Surveillance, 24(4), 1800263.
Schoeman, D., & Fielding, B. C. (2019). Coronavirus envelope protein: Current knowledge. Virology Journal, 16, 69.
https://doi.org/10.1186/s12985-019-1182-0
Otter, J. A., Donskey, C., Yezli, S., Douthwaite, S., Goldenberg, S. D., & Weber, D. J. (2016). Transmission of SARS and MERS coronaviruses and influenza virus in healthcare settings. Journal of Hospital Infection, 92(3), 235–250.
Kampf, G., Todt, D., Pfaender, S., & Steinmann, E. (2020). Persistence of coronaviruses on inanimate surfaces and their inactivation. Journal of Hospital Infection, 104(3), 246–251.
Budden, K. F., Gellatly, S. L., Wood, D. L. A., Cooper, M. A., Morrison, M., Hugenholtz, P., & Hansbro, P. M. (2017). Emerging pathogenic links between microbiota and the gut–lung axis. Nature Reviews Microbiology, 15(1), 55–63.
Dang, A. T., & Marsland, B. J. (2019). Microbiota and the lung immune response. Immunity, 49(2), 192–210.
Ichinohe, T., Pang, I. K., Kumamoto, Y., Peaper, D. R., Ho, J. H., Murray, T. S., & Iwasaki, A. (2011). Microbiota regulates immune defense against respiratory tract influenza A virus infection. Proceedings of the National Academy of Sciences, 108(13), 5354–5359.
Cani, P. D., Amar, J., Iglesias, M. A., Poggi, M., Knauf, C., Bastelica, D., … & Burcelin, R. (2007). Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes, 56(7), 1761–1772.
Bischoff, S. C., Barbara, G., Buurman, W., Ockhuizen, T., Schulzke, J. D., Serino, M., … & Wells, J. M. (2014). Intestinal permeability – a new target for disease prevention and therapy. BMC Gastroenterology, 14, 189.
De Punder, K., & Pruimboom, L. (2015). Stress induces endotoxemia and low-grade inflammation. International Journal of Endocrinology, 2015, 647347.
Trompette, A., Gollwitzer, E. S., Yadava, K., Sichelstiel, A. K., Sprenger, N., Ngom-Bru, C., … & Marsland, B. J. (2014). Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nature Medicine, 20(2), 159–166.
Koh, A., De Vadder, F., Kovatcheva-Datchary, P., & Bäckhed, F. (2016). From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell, 165(6), 1332–1345.
Tan, J., McKenzie, C., Potamitis, M., Thorburn, A. N., Mackay, C. R., & Macia, L. (2014). The role of short-chain fatty acids in health and disease. Advances in Immunology, 121, 91–119.
Hao, Q., Dong, B. R., & Wu, T. (2015). Probiotics for preventing acute upper respiratory tract infections. Cochrane Database of Systematic Reviews, (2), CD006895.
King, S., Glanville, J., Sanders, M. E., Fitzgerald, A., & Varley, D. (2014). Effectiveness of probiotics on the duration of illness in healthy children and adults. British Journal of Nutrition, 112(1), 41–54.
Wang, Y., Li, X., Ge, T., Xiao, Y., Liao, Y., & Cui, Y. (2016). Probiotics for preventing respiratory tract infections in children. European Journal of Clinical Nutrition, 70(4), 494–499.
Furusawa, Y., Obata, Y., Fukuda, S., Endo, T. A., Nakato, G., Takahashi, D., … & Ohno, H. (2013). Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature, 504(7480), 446–450.
Atarashi, K., Tanoue, T., Oshima, K., Suda, W., Nagano, Y., Nishikawa, H., … & Honda, K. (2013). Treg induction by a rationally selected mixture of Clostridia strains. Science, 331(6015), 337–341.
Banat, I. M., Franzetti, A., Gandolfi, I., Bestetti, G., Martinotti, M. G., Fracchia, L., Smyth, T. J., & Marchant, R. (2010). Microbial biosurfactants: Production, applications and future potential. Applied Microbiology and Biotechnology, 87(2), 427–444.
Rodrigues, L. R., Banat, I. M., Teixeira, J. A., & Oliveira, R. (2006). Biosurfactants: Potential applications in medicine. Journal of Antimicrobial Chemotherapy, 57(4), 609–618.
Kok, T., & Nyotohadi, D. (2024). Biosurfactant potential and antiviral activity of multistrain probiotics against SARS-CoV-2. Heliyon, 10(1), e22837.
Zuo, T., Zhang, F., Lui, G. C. Y., Yeoh, Y. K., Li, A. Y. L., Zhan, H., … & Ng, S. C. (2020). Alterations in gut microbiota of patients with COVID-19. Gastroenterology, 159(3), 944–955.
Liu, Q., Mak, J. W. Y., Su, Q., Yeoh, Y. K., Lui, G. C. Y., Ng, S. S. S., … & Chan, F. K. L. (2021). Gut microbiota dynamics in COVID-19. Frontiers in Nutrition, 8, 737773.k
***
*) By: Ge Recta Geson.
Simak breaking news dan berita pilihan TIMES Indonesia langsung dari WhatsApp-mu!
Klik 👉 Channel TIMES Indonesia
Pastikan WhatsApp kamu sudah terpasang.
